Les plantes transgéniques ou plantes génétiquement modifiées Alors que le génie génétique était présenté il y a quelques années comme la panacée à de nombreux problèmes (faim dans le monde, maladies génétiques, cancer, SIDA…), la tendance actuelle, relayée voire amplifiée par les médias, est un rejet global de cette technique alors que nous manquons souvent d’informations et de connaissances nous permettant de nous forger une opinion réfléchie. Définitions La transgénèse permet d’introduire dans des plantes de nouveaux gènes d’abord identifiés, puis isolés et multipliés à volonté. Une fois intégrés dans le génome de la plante, les transgènes sont transmis à la descendance au même titre que tous les autres gènes. On peut ainsi introduire des gènes de résistance, de stérilité, de production de nouvelles protéines etc. Une plante est dite transgénique si un ou plusieurs caractères ont été acquis, non par transfert de gènes entre deux parents lors d’un croisement, mais par des techniques de génie génétique. Les techniques, devenues classiques, de biologie moléculaire permettent de cloner, de séquencer, de découper, de modifier, de recombiner des fragments d’ADN d’origines diverses, le tout pour fabriquer des gènes chimériques. Ces gènes, introduits dans le génome d’une cellule, et si celle-ci donne naissance à un individu, constitueront les transgènes de ce dernier et de sa descendance, puisqu’ils seront transmis comme un caractère génétique classique, dit mendélien. Pour les végétaux comme pour les animaux, si la cible de la transformation n’est pas une cellule, mais un groupe de cellules – un embryon par exemple -, l’individu résultant de la transformation primaire sera une plante chimérique, constituée à la fois de cellules génétiquement transformées et de cellules non modifiées. La régénération d’un individu constitué entièrement de cellules transformées ne pourra se faire que s’il y a sélection des cellules transformées ou que si la transformation ait porté sur le noyau de cellules à l’origine des gamètes. Le succès de toute transgénèse suppose donc la pénétration de l’ADN étranger dans les cellules, son intégration dans le génome, l’aptitude des transgènes à être exprimés, et enfin la possibilité d’obtenir la régénération d’individus entiers à partir des cellules génétiquement modifiées. Techniques de transformations Transfert indirect Agrobacterium tumefaciens et A. rhizogenes sont des bactéries du sol, responsables, l’une de la galle du collet, l’autre de la formation d’un chevelu racinaire au point d’inoculation. Les études menées depuis une vingtaine d’années ont permis de démontrer que ces phénomènes naturels de transformation sont dus à la présence de plasmides Ti (Tumor inducing) ou Ri (Root inducing) dont une partie, nommée ADN-T (ADN Transféré), est transféré par l’agrobactérie au noyau des cellules végétales. Pour que ce transfert d’ADN à la cellule végétale soit possible, il suffit que la séquence d’ADN soit comprise entre les frontières droite et gauche de l’ADN- T et que Agrobacterium qui le porte possède les fonctions dites de virulence, qui permettent la reconnaissance de ses frontières et son transfert vers le noyau des cellules végétales infectées. Les oncogènes de l’ADN-T sont donc remplacés par un gène conférant une résistance à un agent phytotoxique (antibiotique ou herbicide). Parmi les cellules mises en contact par l’agrobactérie modifiée, seules celles qui ont été transformées exprimeront la résistance. Sur un milieu contenant à la fois l’agent de sélection et les hormones de croissance nécessaires à la régénération, seules les cellules transformées seront capables de survivre, de se diviser et de donner naissance à des plantes entières. Transfert direct La plupart des plantes de grande culture sont des monocotylédones, longtemps réputées insensibles à Agrobacterium. En fait, il est maintenant possible d’obtenir du riz transgénique via Agrobacterium ; c’est dans l’établissement de conditions permettant la régénération à partir de cellules somatiques que résident les difficultés. Certaines dicotylédones d’intérêt agronomique, comme le coton ou le soja, s’avèrent également récalcitrantes à la transformation par Agrobacterium. C’est pourquoi différentes techniques de transfert direct ont été mises au point. On a d’abord fait appel à des procédés utilisés avec succès pour transformer des cellules animales. Plus récemment, la biolistique, (méthode mise au point sur des tissus végétaux, qui consiste à bombarder le tissu à transformer de microparticules d’or ou de tungstène recouvertes d’ADN), est venue détrôner les techniques précédentes. Le transfert direct d’ADN ne nécessite pas d’autres techniques que celles permettant d’amplifier et de préparer en quantité l’ADN transféré (gènes d’intérêt et de sélection). Construction de l’ADN transféré L’introduction des transgènes dans le noyau se fait soit par l’intermédiaire d’une bactérie du genre Agrobacterium, soit par des techniques chimiques (électroporation) ou physiques de transfert direct d’ADN. Les transgènes doivent ensuite pouvoir s’exprimer dans les cellules végétales. Qu’ils soient d’origine animale, fongique, bactérienne, ou synthétique, il faut que les signaux permettant leur transcription (ADN vers ARN) et sa régulation soit reconnus par la machinerie cellulaire de l’hôte ; ceux-ci doivent donc être empruntés à des gènes végétaux présentant la même régulation. Pour que la nouvelle protéine soit acheminée vers un compartiment cellulaire particulier, la traduction du messager du gène chimérique devra conduire à une protéine-fusion comportant la séquence de la protéine mature et celle d’un peptide capable de la mener à l’adresse désirée. Arguments pour les OGM Eradication de la faim dans le monde Agriculture propre (OGM résistants aux maladies) Guérison de maladies aujourd’hui incurables Amélioration de la saveur et de la qualité nutritive des aliments Arguments contre les OGM Dissémination risquée et non contrôlée d’espèces modifiées Non-respect du principe de précaution Asservissement des paysans du tiers-monde Absence de traçabilité des OGM Bibliographie Séralini G.-E., 2000. OGM, le vrai débat. Editions Flammarion, 128p.
Tous les articles par _
Elaboration et conservation d’un herbier
Méthodes d’élaboration d’un herbier Principes généraux Après leur détermination et leur identification, les plantes sont séchées sous presse avant d’être fixées par de la colle sans acide sur du papier sans acide. Les planches sont ensuite regroupées par famille en respectant une classification phylogénique. Conseils techniques Séchage. Les plantes sont séchées dans du papier journal qui est changé régulièrement jusqu’au séchage complet (excepté, la feuille directement au contact de la plante). Quand un lot de plantes est épais, il est recommandé de placer quelques planches de carton rigide pour éviter la déformation des plantes.  Pressage. Il ne faut pas que les plantes soient écrasées. La presse peut être simplement constituée de 2 grilles de réfrigérateur maintenues serrées par des sangles ou par différentes masses réparties uniformément sur la surface d’une planche. Qualité des exsiccata. Pour obtenir des échantillons de qualité, le séchage doit commencer le plus rapidement possible après la récolte. Il est souvent constaté que les plantes perdent leur couleur après le séchage, sauf celles contenant naturellement du soufre (exemple : ail). Pour remédier à cet inconvénient, les plantes peuvent être mises en présence de vapeurs soufrées juste après la récolte. Les pastilles de soufre utilisées pour la stérilisation des tonneaux de vins conviennent. Cependant, cette méthode est fastidieuse. Coût d’élaboration La taille des planches est, communément, d’environ 45 cm x 28 cm. Cependant, ces dimensions ne constituent pas un standard obligatoire. Ainsi, Fournier indique dans les Quatre flores de France (1977, réédition) que l’herbier lui-même peut fort bien être constitué sur petit format, par exemple sur des demi-feuilles de papier à machine (à écrire). Les planches que j’utilise mesurent 32 cm x 29 cm et ont un coût unitaire partiel (hors main d’oeuvre) plus de 10 fois inférieur à celui calculé pour les planches traditionnelles, tout en respectant le cahier des charges suivant : papier et colle sans acide, grammage élevé, papier cristal (Bray et al., 1997). La colle utilisée pour fixer les plantes séchées peut être de la colle blanche distribuée avec un pistolet à colle ou encore du rubber cement. Ce dernier produit est vendu sous différentes marques mais aura toujours la même composition chimique, à base de caoutchouc naturel. Etiquetage L’étiquette ou l’enregistrement informatique de chaque planche doit comporter les informations suivantes : – un numéro de référence, – la famille, le genre, l’espèce et les éventuels noms infraspécifiques. Ces indications doivent respecter la nomenclature en vigueur, – l’abréviation du nom de l’auteur spécifique, – le synonyme éventuel, – la date de collecte, – le lieu de collecte. L’utilisation d’un système de positionnement global est recommandé. Il permettra de préciser la latitude et la longitude. Pour l’altitude, compte tenu de l’erreur de l’appareil, il est préférable d’utiliser un altimètre, – le nom du collecteur et/ou le nom de l’identificateur, – la description du biotope ou de l’association phytosociologique (cette dernière indication peut, éventuellement, ne pas être présente sur l’étiquette si elle existe déjà dans la base de données informatisée). Dans le cas d’un premier étiquetage, l’étiquette est placée en bas et à droite. S’il s’agit d’une étiquette apportant des corrections, elle doit être placée en bas et à gauche.
Pressage. Il ne faut pas que les plantes soient écrasées. La presse peut être simplement constituée de 2 grilles de réfrigérateur maintenues serrées par des sangles ou par différentes masses réparties uniformément sur la surface d’une planche. Qualité des exsiccata. Pour obtenir des échantillons de qualité, le séchage doit commencer le plus rapidement possible après la récolte. Il est souvent constaté que les plantes perdent leur couleur après le séchage, sauf celles contenant naturellement du soufre (exemple : ail). Pour remédier à cet inconvénient, les plantes peuvent être mises en présence de vapeurs soufrées juste après la récolte. Les pastilles de soufre utilisées pour la stérilisation des tonneaux de vins conviennent. Cependant, cette méthode est fastidieuse. Coût d’élaboration La taille des planches est, communément, d’environ 45 cm x 28 cm. Cependant, ces dimensions ne constituent pas un standard obligatoire. Ainsi, Fournier indique dans les Quatre flores de France (1977, réédition) que l’herbier lui-même peut fort bien être constitué sur petit format, par exemple sur des demi-feuilles de papier à machine (à écrire). Les planches que j’utilise mesurent 32 cm x 29 cm et ont un coût unitaire partiel (hors main d’oeuvre) plus de 10 fois inférieur à celui calculé pour les planches traditionnelles, tout en respectant le cahier des charges suivant : papier et colle sans acide, grammage élevé, papier cristal (Bray et al., 1997). La colle utilisée pour fixer les plantes séchées peut être de la colle blanche distribuée avec un pistolet à colle ou encore du rubber cement. Ce dernier produit est vendu sous différentes marques mais aura toujours la même composition chimique, à base de caoutchouc naturel. Etiquetage L’étiquette ou l’enregistrement informatique de chaque planche doit comporter les informations suivantes : – un numéro de référence, – la famille, le genre, l’espèce et les éventuels noms infraspécifiques. Ces indications doivent respecter la nomenclature en vigueur, – l’abréviation du nom de l’auteur spécifique, – le synonyme éventuel, – la date de collecte, – le lieu de collecte. L’utilisation d’un système de positionnement global est recommandé. Il permettra de préciser la latitude et la longitude. Pour l’altitude, compte tenu de l’erreur de l’appareil, il est préférable d’utiliser un altimètre, – le nom du collecteur et/ou le nom de l’identificateur, – la description du biotope ou de l’association phytosociologique (cette dernière indication peut, éventuellement, ne pas être présente sur l’étiquette si elle existe déjà dans la base de données informatisée). Dans le cas d’un premier étiquetage, l’étiquette est placée en bas et à droite. S’il s’agit d’une étiquette apportant des corrections, elle doit être placée en bas et à gauche.
Relations entre les organismes
Relations entre les organismes Réseau trophique Des chaînes alimentaires sont formées entre différentes espèces : plantes vertes (producteurs) ; herbivores (consommateurs primaires) ; petits et grands carnivores, nécrophages (consommateurs secondaires). Les êtres n’ayant pas de régime alimentaire spécialisé (omnivores par exemple) établissent des relations entre les différentes chaînes alimentaires qui, combinées entre elles, forment un réseau trophique. Un réseau trophique peut englober différentes biocénoses[1] ou écosystèmes[2]. Certaines espèces sont particulièrement difficiles à classer dans une biocénose : les larves d’amphibiens sont dans un milieu aquatique alors que les adultes sont terrestres. Equilibre biologique La biocénose est un ensemble supra-individuel et supra-spécifique possédant des capacités de régulation : si un hiver rigoureux a détruit beaucoup de larves d’insectes, les insectivores seront moins nombreux….Dans la pratique, les valeurs des populations varient autour d’une valeur moyenne appelée équilibre biologique. Plus le réseau trophique est complexe plus les variations seront faibles (bien que les variations d’une seule espèce auront des impacts sur un plus grand nombre d’espèces). Principes biocénotiques fondamentaux Plus les conditions de vie d’un biotope peuvent varier, plus le nombre d’espèces est élevé. Par exemple dans les régions d’estuaire, il est possible de trouver des poissons d’eau douce et d’eau salée. Quand les conditions de vie s’écartent d’un minimum vital (chaleur, ressources en eau), plus le nombre d’espèces est faible et plus le nombre d’individus par espèce est important. Par exemple, le nombre d’espèces de la strate arborée va en décroissant de la forêt équatoriale, à celle tropicale, puis tempérée et sahélienne. Les biocénoses ayant un très grand nombre d’espèces ont des populations relativement stables dont la survie est moins difficile (hors intervention humaine). A l’inverse, des biocénoses aux espèces peu nombreuses peuvent subir des variations de populations très importantes : en zone sahélienne, la sécheresse peut avoir des effets désastreux. Cycle alimentaire Dans une chaîne alimentaire, trois catégories sont distinguées : – les producteurs sont les végétaux autotrophes qui transforment les substances minérales en substances organiques, – les consommateurs (végétaux hétérotrophes, animaux) se nourissent directement ou indirectement des matières organiques élaborées par les producteurs, – les décomposeurs (principalement des bactéries) qui réduisent les cadavres, les déchets des végétaux et des animaux en des substances minérales pouvant être à nouveau disponibles pour les producteurs. ——————————————————————————– [1] biocénose : ensemble d’êtres vivants qui se constitue dans des des conditions écologiques données et qui se maintient dans un équilibre dynamique. [2] écosystème : une biocénose avec l’ensemble des facteurs inorganiques du biotope qu’elle occupe constitue un écosystème.
Banque de semences : outil de conservation
Banque de semences Catégories de semences Une semence contient un embryon différencié (ébauche de racine et de tige, un ou deux cotylédons[1] qui sont des feuilles embryonnaires), des réserves situées dans les cotylédons ou dans des tissus spécialisés (albumen[2] ou périsperme[3] chez les Angiospermes, endosperme[4] chez les Gymnospermes) (Côme, 1970 & 1992). Deux catégories de semences peuvent être considérées (Côme, 1992) : d’une part, des semences orthodoxes à faible teneur en eau, supportant la dessiccation et une conservation au froid, d’autre part, des semences récalcitrantes à la dessiccation, à teneur en eau élevée, ne supportant pas, en général, une conservation au froid. 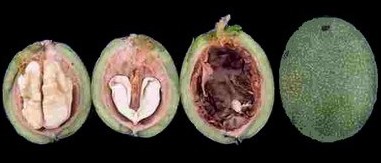 Photo de noix : La semence du fruit de noyer est récalcitrante et ne peut pas être conservée au froid. Une autre catégorie dite de “ semences intermédiaires ” peut, éventuellement, être prise en compte : les semences supportent une dessiccation et une conservation au froid dans des conditions strictement contrôlées et pour une période plus courte que pour les semences orthodoxes (Hong et al, 1996). Les deux catégories, orthodoxe et récalcitrante, sont importantes car elles sont liées au mode de conservation et à la longévité des semences. Ainsi, les semences orthodoxes peuvent être placées au congélateur de -18°C à -33°C, pour une conservation à long terme, si leur teneur en eau est maintenue la plus basse possible. Au contraire, 15°C est un seuil à ne pas franchir pour conserver, à moyen ou long terme, la majorité des semences récalcitrantes (Cromarty et al, 1990). Si la teneur en eau permet de déterminer la catégories des semences, Hong et al. (1996) ont aussi prouvé les corrélations existant entre ces catégories et le poids ou le volume des semences. En résumé, plus une semence est petite et/ou dense plus la probabilité pour qu’elle soit orthodoxe est élevée. Collecte et stockage Les semences sont collectées soit en milieu naturel, soit dans des parcelles cultivées. Pour ces dernières récoltes, les pieds-mères sont sélectionnés pour que la probabilité d’hybridation inter- et intra-spécifique non contrôlée soit la plus faible possible. La probabilité d’hybridation spontanée inter-générique étant suffisamment faible pour être considérée comme nulle, le critère de sélection est l’absence ou l’éloignement important, sur le site d’une autre espèce du même genre. Cependant, 3 exceptions à cette règle sont prises en compte. La première permet de considérer le risque d’hybridation comme faible quand les pieds d’espèces différentes sont très éloignés sur le même site. Cette exception à la règle générale se justifie d’autant plus pour des espèces entomophiles introduites. En effet, dans ce cas, les insectes ne sont pas spécialisés et chaque pollinisation sera un événement aléatoire et indépendant : si n est le nombre total de taxons sur le site, la probabilité d’hybridation peut être évaluée, en première approximation, à n-2 . La 2ème exception considère qu’il ne peut y avoir d’hybridation entre deux espèces du même genre si les périodes de floraison sont bien distinctes. Enfin, la 3ème exception s’applique quand la pratique a déjà prouvé que les semis de certains taxons conservaient les caractères des pieds-mères. Les semences orthodoxes sont conservées à l’obscurité, dans des boites hermétiques, en présence de gel de silice, à 5°C ou, éventuellement, au congélateur à -18°C ou -33°C. Les semences récalcitrantes sont conservées à l’obscurité, à 15°C. Références bibliographiques Côme D., 1970. Les obstacles à la Germination. Masson & Cie ed., Paris : 162 pp. Côme D., Corbineau F., 1992. Les végétaux et le froid. Dans les semences et le froid, Côme pub., Hermann ed., Paris : 401-461. Cromarty A. S., Ellis R. H., Roberts E.H., 1990. The Design of Seed Storage Facilities for Genetic Conservation. Edition révisée. International Board for Plant Genetic Resources, Rome : 100 pp. Hong T. D., Linington S., Ellis R. H., 1996. Compendium of Information on Seed Storage Behaviour. International Plant Genetic Resources Institute, Rome : sous presse. Young J. A., Young C. G., 1992. Seeds of woody plants in North America. Revised and enlarged edition. Dioscorides Press, Portland : 407 pp. ——————————————————————————– [1] Respectivement, classe des Monocotyledonae et celle des Dicotyledonae. [2] Tissu triploïde issu de la fécondation par un spermatozoïde des deux noyaux accessoires du sac embryonnaire. [3] Tissu diploïde d’origine maternelle, nucelle chargé de réserves [4] Tissu haploïde coenocytique, gamétophyte femelle.
Photo de noix : La semence du fruit de noyer est récalcitrante et ne peut pas être conservée au froid. Une autre catégorie dite de “ semences intermédiaires ” peut, éventuellement, être prise en compte : les semences supportent une dessiccation et une conservation au froid dans des conditions strictement contrôlées et pour une période plus courte que pour les semences orthodoxes (Hong et al, 1996). Les deux catégories, orthodoxe et récalcitrante, sont importantes car elles sont liées au mode de conservation et à la longévité des semences. Ainsi, les semences orthodoxes peuvent être placées au congélateur de -18°C à -33°C, pour une conservation à long terme, si leur teneur en eau est maintenue la plus basse possible. Au contraire, 15°C est un seuil à ne pas franchir pour conserver, à moyen ou long terme, la majorité des semences récalcitrantes (Cromarty et al, 1990). Si la teneur en eau permet de déterminer la catégories des semences, Hong et al. (1996) ont aussi prouvé les corrélations existant entre ces catégories et le poids ou le volume des semences. En résumé, plus une semence est petite et/ou dense plus la probabilité pour qu’elle soit orthodoxe est élevée. Collecte et stockage Les semences sont collectées soit en milieu naturel, soit dans des parcelles cultivées. Pour ces dernières récoltes, les pieds-mères sont sélectionnés pour que la probabilité d’hybridation inter- et intra-spécifique non contrôlée soit la plus faible possible. La probabilité d’hybridation spontanée inter-générique étant suffisamment faible pour être considérée comme nulle, le critère de sélection est l’absence ou l’éloignement important, sur le site d’une autre espèce du même genre. Cependant, 3 exceptions à cette règle sont prises en compte. La première permet de considérer le risque d’hybridation comme faible quand les pieds d’espèces différentes sont très éloignés sur le même site. Cette exception à la règle générale se justifie d’autant plus pour des espèces entomophiles introduites. En effet, dans ce cas, les insectes ne sont pas spécialisés et chaque pollinisation sera un événement aléatoire et indépendant : si n est le nombre total de taxons sur le site, la probabilité d’hybridation peut être évaluée, en première approximation, à n-2 . La 2ème exception considère qu’il ne peut y avoir d’hybridation entre deux espèces du même genre si les périodes de floraison sont bien distinctes. Enfin, la 3ème exception s’applique quand la pratique a déjà prouvé que les semis de certains taxons conservaient les caractères des pieds-mères. Les semences orthodoxes sont conservées à l’obscurité, dans des boites hermétiques, en présence de gel de silice, à 5°C ou, éventuellement, au congélateur à -18°C ou -33°C. Les semences récalcitrantes sont conservées à l’obscurité, à 15°C. Références bibliographiques Côme D., 1970. Les obstacles à la Germination. Masson & Cie ed., Paris : 162 pp. Côme D., Corbineau F., 1992. Les végétaux et le froid. Dans les semences et le froid, Côme pub., Hermann ed., Paris : 401-461. Cromarty A. S., Ellis R. H., Roberts E.H., 1990. The Design of Seed Storage Facilities for Genetic Conservation. Edition révisée. International Board for Plant Genetic Resources, Rome : 100 pp. Hong T. D., Linington S., Ellis R. H., 1996. Compendium of Information on Seed Storage Behaviour. International Plant Genetic Resources Institute, Rome : sous presse. Young J. A., Young C. G., 1992. Seeds of woody plants in North America. Revised and enlarged edition. Dioscorides Press, Portland : 407 pp. ——————————————————————————– [1] Respectivement, classe des Monocotyledonae et celle des Dicotyledonae. [2] Tissu triploïde issu de la fécondation par un spermatozoïde des deux noyaux accessoires du sac embryonnaire. [3] Tissu diploïde d’origine maternelle, nucelle chargé de réserves [4] Tissu haploïde coenocytique, gamétophyte femelle.
