Chaque faisceau cribro-vasculaire est constitué comme suit : – le xylème qui conduit la sève brute (eaux et éléments minéraux en provenance des racines) est situé sur la face intérieure du faisceau, la plus proche de l’axe de la tige. – le phloème qui conduit la sève élaborée (eau et sucres formés dans les feuilles par la photosynthèse) est situé sur la face externe du faisceau, la plus éloignée de l’axe de la tige. Les éléments conducteurs, les tubes criblés pour le phloème (sève élaborée) et les vaisseaux du xylème (sève brute) ont donné leur nom à l’ensemble : les faisceaux cribro-vasculaires. Le xylème et le phloème sont séparés par une bande de cellules non différenciées (encore embryonnaires) dont le fonctionnement est bloqué : elles constituent le cambium fasciculaire. Elles ne reprendront leur activité que durant l’élaboration des structures secondaires. Dans les rayons médullaires, certaines cellules embryonnaires inactives sont prêtes à reprendre leur activité de multiplication ; elles constituent le cambium inter-fasciculaire. Les tissus conducteurs peuvent être entourés d’un anneau de cellules lignifiées (contenant de la lignine, un composant chimique durcissant les cellules) appelées sclérenchyme. Cependant, le sclérenchyme est alors incomplet sur les 2 côtés du faisceau cribro-vasculaire au niveau du cambium, ce qui rendra possible sa reprise d’activité au moment de l’élaboration des structures secondaires. Le faisceau est dit collatéral ouvert. Chez certaines plantes (Cucurbitaceae par exemple), le phloème est présent de part et d’autre du xylème, il s’agit d’un faisceau bicollatéral ouvert.
Archives par mot-clé : anatomie
Structure primaire de la racine
En section transversale, la racine est divisée en 2 parties. Comme dans la tige, la plus externe est l’écorce. Elle est cependant plus épaisse. En périphérie, elle est limitée par l’épiderme et vers le centre par l’endoderme, absent dans la tige, qui constitue une limite nette avec la partie plus interne. La partie plus interne de la racine est le cylindre central qui contient le péricycle, les tissus conducteurs et la moelle. Contrairement à la tige, le xylème et le phloème ne sont pas groupés en faisceaux : ils sont placés sur des rayons différents et alternes dans un cylindre central dont la limite périphérique est le péricycle qui se trouve au contact de l’endoderme. 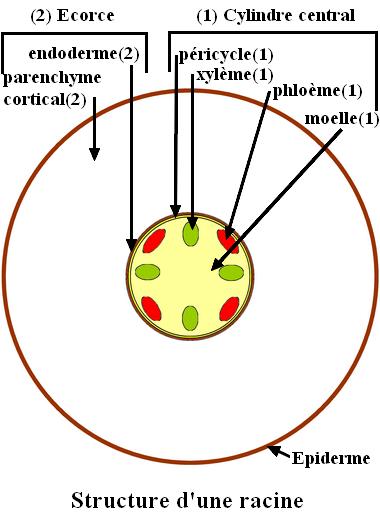 Le péricycle est constitué d’une seule couche de cellules. Il fonctionne comme un méristème secondaire et forme des racines latérales. La moelle qui est importante dans une structure primaire de tige est très réduite dans une racine, voire absente dans les cas où les rayons de xylème se rejoignent au centre du cylindre central.
Le péricycle est constitué d’une seule couche de cellules. Il fonctionne comme un méristème secondaire et forme des racines latérales. La moelle qui est importante dans une structure primaire de tige est très réduite dans une racine, voire absente dans les cas où les rayons de xylème se rejoignent au centre du cylindre central.
Croissance en longueur de la racine
En section longitudinale, les différentes zones de la racine sont les suivantes : – la coiffe à l’extrémité de la racine. Les cellules les plus internes renouvellent au fur et à mesure les cellules les plus externes, qui disparaissent au contact du sol. – le méristème apical ou point végétatif contenant au dessus de la zone quiescente des cellules embryonnaires se divisant. Placé à l’abri de la coiffe, il fournit les cellules nécessaires à l’allongement de la racine. – la zone de croissance et de différenciation au dessus du point végétatif. – la zone pilifère où les cellules épidermiques développent des poils absorbant les éléments minéraux du sol. – la zone de ramification où les poils absorbants sont absents mais où des racines secondaires peuvent être formées par le péricycle.
Structure de la tige des Monocotylédones.
Dans une tige de Monocotylédone, les faisceaux cribro-vasculaires sont disposés en plusieurs cercles concentriques. De plus, le cambium fasciculaire ou inter-fasciculaire est absent. La moelle est réduite ou absente. 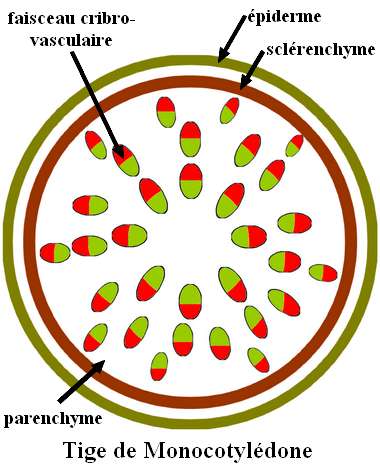 Le sclérenchyme peut être présent sous forme d’un cercle entourant l’ensemble des faisceaux cribro-vasculaires (cas de l’Asperge) ou engainant chacun de ces faisceaux. Dans ce dernier cas, la gaine de sclérenchyme ferme entièrement le faisceau cribro-vasculaire alors que chez les Dicotylédones, elle est discontinue au niveau du cambium. On dit que le faisceau est collatéral fermé chez les Monocotylédones alors qu’il est collatéral ouvert chez les Dicotylédones.
Le sclérenchyme peut être présent sous forme d’un cercle entourant l’ensemble des faisceaux cribro-vasculaires (cas de l’Asperge) ou engainant chacun de ces faisceaux. Dans ce dernier cas, la gaine de sclérenchyme ferme entièrement le faisceau cribro-vasculaire alors que chez les Dicotylédones, elle est discontinue au niveau du cambium. On dit que le faisceau est collatéral fermé chez les Monocotylédones alors qu’il est collatéral ouvert chez les Dicotylédones.
Cernes de croissance, coeur et aubier du bois
L’activité du cambium est saisonnière. Les divisions de cellules s’arrêtent en hiver, reprennent au printemps avec la formation de vaisseaux de diamètre important. Cette saisonnalité se traduit par des cernes annuels de croissance permettant de dater l’âge du bois. La sève n’est souvent conduite que dans les cernes les plus récents. Dans les cernes les plus profonds, le bois durcit chez certains arbres. La zone centrale appelée cœur accumulant des tanins contre les champignons responsables du pourrissement. Ces tanins sont de couleur foncée : noire dans le bois d’ébène. La partie périphérique du bois, plus claire, s’appelle l’aubier. Quand le cœur du bois n’est pas durci comme chez le Saule, le Peuplier ou le Tilleul, des champignons provoquent sa pourriture et le creuse.