Certaines plantes n’ont pas de vraies fleurs, c’est le cas des pins et des sapins. Les ovules ne sont pas protégés dans un ovaire et sont placés nus sous une écaille d’où leur nom Gymnospermes qui signifie graines nues. Il n’y a ni pétale ni sépale. L’ovule après la fécondation donnera bien une graine mais il n’y aura pas de fruit. D’autres plantes n’auront ni fleur ni graine, c’est le cas des fougères ou des prêles. Il n’y a pas de survie de l’espèce sous forme de graine et l’embryon devra avoir les conditions nécessaires à son développement juste après la fécondation sinon il mourra. Pour ces raisons, la répartition de ces plantes est souvent limitée à des zones humides.
Archives par mot-clé : fruit
Les plantes à fleurs
De l’extérieur vers l’intérieur, les fleurs sont composées : – des sépales, généralement verts dont l’ensemble est appelé calice, – des pétales, généralement colorés dont l’ensemble est appelé corolle, – des étamines, organes reproducteurs mâles produisant le pollen, – des carpelles, organes élémentaires reproducteurs femelles dont l’ensemble est appelé pistil. Les sépales et les pétales sont généralement au nombre de 3 (lis), 4 (chou, giroflée) ou 5 (renoncule, fraisier, pomme de terre), parfois plus. Quand les sépales et les pétales se ressemblent, ils sont appelés tépales. Ils sont absents chez certaines plantes (miscanthus, bambou). Les pétales peuvent être libres entre eux (giroflée, rose, tulipe, pois) ou soudés (muflier, digitale, orchidée). La fleur peut avoir une forme de cercle, elle est alors régulière (giroflée) ; ou une symétrie plane, elle alors irrégulière (pois, orchidée). Les fleurs sont portées par un pédoncule floral. A la base du pédoncule se trouve les bractées qui ressemblent à de petites feuilles. Les bractées sont normalement vertes mais sont parfois vivement colorées (poinsettia). Les fleurs peuvent être solitaires ou plus nombreuses et disposées en inflorescence. Parfois, elles sont petites et regroupées ensemble en tête à l’extrémité du pédoncule floral. Par exemple, la marguerite n’est pas constituée d’une seule fleur mais de plusieurs : des fleurs blanches en périphérie et en forme de langue, et des fleurs jaunes au centre en forme de tube. Les étamines ont une partie fine en forme de fil, le filet qui porte à son extrémité des sacs, les anthères, qui libéreront le pollen au moment de la pollinisation. Parfois, les étamines sont absentes et la fleur est unisexuée et femelle. Parfois, les carpelles sont absents et la fleur est mâle. Les fleurs unisexuées peuvent être portées sur une seule plante (bégonia) ou sur des plantes différentes (kiwi). A sa base, le carpelle a une partie ronde, l’ovaire qui contient l’ovule. A son extrémité, le stigmate est l’endroit collant où se dépose le pollen. Entre l’ovaire et le stigmate, se trouve une partie intermédiaire plus fine, le style qui est plus ou moins long. Le pollen donnera le spermatozoïde qui fécondera l’ovule. L’ovule fécondé par le spermatozoïde, donnera l’embryon qui se trouvera protégé dans la graine. L’ovaire deviendra le fruit qui protégera la graine elle-même et facilitera sa dispersion. Normalement, le spermatozoïde ne contient qu’un seul stock de chromosomes comme l’ovule. L’embryon aura donc 2 stocks de chromosomes, celui de l’ovule et celui du spermatozoïde. Cet embryon donnera une plante dite diploïde. Chez certaines plantes, le stock de chromosomes, toujours pair sera beaucoup plus élevé (plantes polyploïdes). Ce phénomène existe dans la nature mais est relativement rare. Les processus de sélection favorisent la présence de plantes ayant un plus grand nombre de chromosomes. En horticulture, cette augmentation du nombre de chromosomes est souvent liée à l’augmentation de la taille de la fleur ou du nombre de pétales. En agriculture, les plantes polyploïdes produiront plus de protéines ou de glucides de réserves. Ainsi, le blé cultivé pour la fabrication du pain contient 42 stocks de chromosomes alors que ses parents sauvages n’en possèdent que 2.
Banque de semences : outil de conservation
Banque de semences Catégories de semences Une semence contient un embryon différencié (ébauche de racine et de tige, un ou deux cotylédons[1] qui sont des feuilles embryonnaires), des réserves situées dans les cotylédons ou dans des tissus spécialisés (albumen[2] ou périsperme[3] chez les Angiospermes, endosperme[4] chez les Gymnospermes) (Côme, 1970 & 1992). Deux catégories de semences peuvent être considérées (Côme, 1992) : d’une part, des semences orthodoxes à faible teneur en eau, supportant la dessiccation et une conservation au froid, d’autre part, des semences récalcitrantes à la dessiccation, à teneur en eau élevée, ne supportant pas, en général, une conservation au froid. 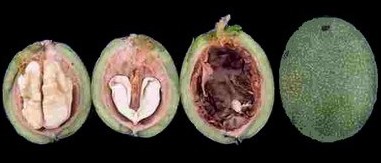 Photo de noix : La semence du fruit de noyer est récalcitrante et ne peut pas être conservée au froid. Une autre catégorie dite de “ semences intermédiaires ” peut, éventuellement, être prise en compte : les semences supportent une dessiccation et une conservation au froid dans des conditions strictement contrôlées et pour une période plus courte que pour les semences orthodoxes (Hong et al, 1996). Les deux catégories, orthodoxe et récalcitrante, sont importantes car elles sont liées au mode de conservation et à la longévité des semences. Ainsi, les semences orthodoxes peuvent être placées au congélateur de -18°C à -33°C, pour une conservation à long terme, si leur teneur en eau est maintenue la plus basse possible. Au contraire, 15°C est un seuil à ne pas franchir pour conserver, à moyen ou long terme, la majorité des semences récalcitrantes (Cromarty et al, 1990). Si la teneur en eau permet de déterminer la catégories des semences, Hong et al. (1996) ont aussi prouvé les corrélations existant entre ces catégories et le poids ou le volume des semences. En résumé, plus une semence est petite et/ou dense plus la probabilité pour qu’elle soit orthodoxe est élevée. Collecte et stockage Les semences sont collectées soit en milieu naturel, soit dans des parcelles cultivées. Pour ces dernières récoltes, les pieds-mères sont sélectionnés pour que la probabilité d’hybridation inter- et intra-spécifique non contrôlée soit la plus faible possible. La probabilité d’hybridation spontanée inter-générique étant suffisamment faible pour être considérée comme nulle, le critère de sélection est l’absence ou l’éloignement important, sur le site d’une autre espèce du même genre. Cependant, 3 exceptions à cette règle sont prises en compte. La première permet de considérer le risque d’hybridation comme faible quand les pieds d’espèces différentes sont très éloignés sur le même site. Cette exception à la règle générale se justifie d’autant plus pour des espèces entomophiles introduites. En effet, dans ce cas, les insectes ne sont pas spécialisés et chaque pollinisation sera un événement aléatoire et indépendant : si n est le nombre total de taxons sur le site, la probabilité d’hybridation peut être évaluée, en première approximation, à n-2 . La 2ème exception considère qu’il ne peut y avoir d’hybridation entre deux espèces du même genre si les périodes de floraison sont bien distinctes. Enfin, la 3ème exception s’applique quand la pratique a déjà prouvé que les semis de certains taxons conservaient les caractères des pieds-mères. Les semences orthodoxes sont conservées à l’obscurité, dans des boites hermétiques, en présence de gel de silice, à 5°C ou, éventuellement, au congélateur à -18°C ou -33°C. Les semences récalcitrantes sont conservées à l’obscurité, à 15°C. Références bibliographiques Côme D., 1970. Les obstacles à la Germination. Masson & Cie ed., Paris : 162 pp. Côme D., Corbineau F., 1992. Les végétaux et le froid. Dans les semences et le froid, Côme pub., Hermann ed., Paris : 401-461. Cromarty A. S., Ellis R. H., Roberts E.H., 1990. The Design of Seed Storage Facilities for Genetic Conservation. Edition révisée. International Board for Plant Genetic Resources, Rome : 100 pp. Hong T. D., Linington S., Ellis R. H., 1996. Compendium of Information on Seed Storage Behaviour. International Plant Genetic Resources Institute, Rome : sous presse. Young J. A., Young C. G., 1992. Seeds of woody plants in North America. Revised and enlarged edition. Dioscorides Press, Portland : 407 pp. ——————————————————————————– [1] Respectivement, classe des Monocotyledonae et celle des Dicotyledonae. [2] Tissu triploïde issu de la fécondation par un spermatozoïde des deux noyaux accessoires du sac embryonnaire. [3] Tissu diploïde d’origine maternelle, nucelle chargé de réserves [4] Tissu haploïde coenocytique, gamétophyte femelle.
Photo de noix : La semence du fruit de noyer est récalcitrante et ne peut pas être conservée au froid. Une autre catégorie dite de “ semences intermédiaires ” peut, éventuellement, être prise en compte : les semences supportent une dessiccation et une conservation au froid dans des conditions strictement contrôlées et pour une période plus courte que pour les semences orthodoxes (Hong et al, 1996). Les deux catégories, orthodoxe et récalcitrante, sont importantes car elles sont liées au mode de conservation et à la longévité des semences. Ainsi, les semences orthodoxes peuvent être placées au congélateur de -18°C à -33°C, pour une conservation à long terme, si leur teneur en eau est maintenue la plus basse possible. Au contraire, 15°C est un seuil à ne pas franchir pour conserver, à moyen ou long terme, la majorité des semences récalcitrantes (Cromarty et al, 1990). Si la teneur en eau permet de déterminer la catégories des semences, Hong et al. (1996) ont aussi prouvé les corrélations existant entre ces catégories et le poids ou le volume des semences. En résumé, plus une semence est petite et/ou dense plus la probabilité pour qu’elle soit orthodoxe est élevée. Collecte et stockage Les semences sont collectées soit en milieu naturel, soit dans des parcelles cultivées. Pour ces dernières récoltes, les pieds-mères sont sélectionnés pour que la probabilité d’hybridation inter- et intra-spécifique non contrôlée soit la plus faible possible. La probabilité d’hybridation spontanée inter-générique étant suffisamment faible pour être considérée comme nulle, le critère de sélection est l’absence ou l’éloignement important, sur le site d’une autre espèce du même genre. Cependant, 3 exceptions à cette règle sont prises en compte. La première permet de considérer le risque d’hybridation comme faible quand les pieds d’espèces différentes sont très éloignés sur le même site. Cette exception à la règle générale se justifie d’autant plus pour des espèces entomophiles introduites. En effet, dans ce cas, les insectes ne sont pas spécialisés et chaque pollinisation sera un événement aléatoire et indépendant : si n est le nombre total de taxons sur le site, la probabilité d’hybridation peut être évaluée, en première approximation, à n-2 . La 2ème exception considère qu’il ne peut y avoir d’hybridation entre deux espèces du même genre si les périodes de floraison sont bien distinctes. Enfin, la 3ème exception s’applique quand la pratique a déjà prouvé que les semis de certains taxons conservaient les caractères des pieds-mères. Les semences orthodoxes sont conservées à l’obscurité, dans des boites hermétiques, en présence de gel de silice, à 5°C ou, éventuellement, au congélateur à -18°C ou -33°C. Les semences récalcitrantes sont conservées à l’obscurité, à 15°C. Références bibliographiques Côme D., 1970. Les obstacles à la Germination. Masson & Cie ed., Paris : 162 pp. Côme D., Corbineau F., 1992. Les végétaux et le froid. Dans les semences et le froid, Côme pub., Hermann ed., Paris : 401-461. Cromarty A. S., Ellis R. H., Roberts E.H., 1990. The Design of Seed Storage Facilities for Genetic Conservation. Edition révisée. International Board for Plant Genetic Resources, Rome : 100 pp. Hong T. D., Linington S., Ellis R. H., 1996. Compendium of Information on Seed Storage Behaviour. International Plant Genetic Resources Institute, Rome : sous presse. Young J. A., Young C. G., 1992. Seeds of woody plants in North America. Revised and enlarged edition. Dioscorides Press, Portland : 407 pp. ——————————————————————————– [1] Respectivement, classe des Monocotyledonae et celle des Dicotyledonae. [2] Tissu triploïde issu de la fécondation par un spermatozoïde des deux noyaux accessoires du sac embryonnaire. [3] Tissu diploïde d’origine maternelle, nucelle chargé de réserves [4] Tissu haploïde coenocytique, gamétophyte femelle.